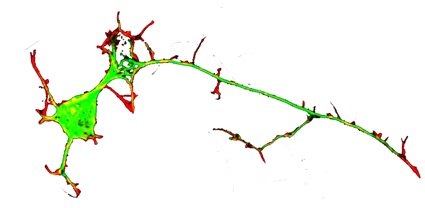
Axonal structure An axon is a cylindrical process that extends to three feet or longer (in humans) from a neuronal cell body, transmitting electrical signals to a neuron’s synaptic targets. Axons may be unbranched or may branch infrequently with more frequent branches at the terminal. Axonal diameters can be less than a micron or as wide as 20 microns. Axonal structure includes an encompassing plasma membrane as an outer skin with many internal mitochondria, and diverse vesicular organelles, all supported by a complex cytoskeleton of longitudinally oriented microtubules and neurofilaments and a network of actin filaments attached to the plasma membrane (40). Although axons of large motor neurons contain >99% of the cell’s volume, in most cases most axonal components are synthesized in the cell body. Nevertheless, axonally synthesized components have critical roles in maintaining axonal function and in localized physiological adaptations, such as learning and memory. Axonal materials are transported from the cell body to terminals and back by motor molecules that bind to cargo and move it along rail-like microtubule arrays at rates that vary from one to several hundred mm/day.
Axonal formation In developing nervous systems neurons first sprout immature processes called neurites, of which one matures into an axon. Neurites arise from sites at the neuronal perimeter that are initially marked by protrusion of finger-like filopodia, which are dynamic elaborations of the actin filament network. Two key events are required to sprout a neurite. One is that a filopodium adheres to adjacent extracellular structures or other surfaces. The other key event is that microtubules advance into an adherent filopodium. Two motor proteins provide “push” and “pull” to direct microtubules into a filopodium. Cytoplasmic dynein motors “push” microtubules toward a filopodium and myosin II motors interact with filopodial actin filaments to “pull” microtubules into a filopodium. Microtubules are dynamic polymers, that grow and shrink by adding or losing subunits at the distal ends. Once a microtubule tip enters a filopodium, the microtubule can advance further by growing its distal tip or by moving forward.
Axonal elongation Once a nascent neurite with microtubules and a motile adherent tip (that is, a growth cone) has formed, the neurite can grow, as long as (1) filopodial protrusion and adhesion continue, (2) microtubules advance down the elongating neurite, and (3) the plasma membrane expands. Short microtubules move into a neurite via dynein motors and lengthen by tubulin monomers adding to their distal ends. After lengthening further, individual microtubules become immobile, so optimal neurite growth involves continual microtubule lengthening by subunit addition, especially at the distal neurite tip (5,13,15). The plasma membrane expands by intercalation of protein and lipid components at many points along a growing neurite, although more addition occurs at the distal growth cone.
Axonal branching Axonal (or neurite) branching occurs along an axonal shaft in a similar manner to neurite formation at the cell body, that is, a local site of protrusion and adhesion must arise, and a microtubule tip(s) must be directed into a nascent branch (25,26). Branches also form at growth cones via local segregation of a portion of the protrusive activity and microtubule tips that are abundantly present in growth cones (6, 8).